通常我们拿到了肿瘤相关的单细胞转录组的表达量矩阵后的第一层次降维聚类分群通常是:
- immune (CD45+,PTPRC),
- epithelial/cancer (EpCAM+,EPCAM),
- stromal (CD10+,MME,fibro or CD31+,PECAM1,endo)
参考我前面介绍过 CNS图表复现08—肿瘤单细胞数据第一次分群通用规则,这3大单细胞亚群构成了肿瘤免疫微环境的复杂。绝大部分文章都是抓住免疫细胞亚群进行细分,包括淋巴系(T,B,NK细胞)和髓系(单核,树突,巨噬,粒细胞)的两大类作为第二次细分亚群。但是也有不少文章是抓住stromal 里面的 fibro 和endo进行细分,并且编造生物学故事的。
前面我们已经介绍了心肝脾肺肾等多个器官的上皮细胞的细分亚群, 以及免疫细胞里面的髓系和B细胞细分亚群:
其中髓系的(单核,树突,巨噬,粒细胞)的细分亚群,主要是树突细胞细分亚群比较多,是cDC1,cDC2,cDC3,以及pDC。然后是单核就区分成为了:
- Mono_CD14 , classical CD14+CD16-
- Mono_FCGR3A ,non-classical CD14-CD16+ monocytes,
当然了,上面的CD14和CD16的单核细胞仍然是可以继续细分的,而且也确实有很多文献做了细分。
也就是说每个大亚群都可以内部细分亚群,然后继续细分,子子孙孙无穷尽。不过实际情况下,大家一般来说区分到第四层级即可,比如Treg属于cd4的T细胞,然后属于T细胞,属于淋巴细胞,属于免疫细胞!
标准的细胞谱系发育层级命名
比如2021年11月发表在《cancer cell杂志》的文献:《Signatures of plasticity, metastasis, and immunosuppression in an atlas of human small cell lung cancer》就提到了:All scRNA-seq data were merged, normalized, batch-cor- rected, and clustered to identify coarse cell types, including epithelial, mesenchymal, lymphoid, and myeloid cells ,也就是说 上皮细胞,基质细胞,淋巴系和髓系。然后是其中上皮细胞继续细分:
- Absorptive and Secretory subtrees.
- The Absorptive subtree included further subtrees for Transit Amplifying (TA) cells (Absorptive TA 1, Absorptive TA 2), Immature cells (Immature Enterocytes 1, Immature Enterocytes 2, Enterocyte Progenitors), and Mature cells (Enterocytes, BEST4+ Enterocytes).
- The Secretory subtree included subtrees for progenitor cells (Secretory TA, Immature Goblet) and for mature cells (Goblet, Tuft, and Enteroendocrine).
- Stem cells, Cycling TA cells, and M cells were placed directly under the node corresponding to all epithelial cells.
而 Stromal subtree had Fibroblast, Endothelial, and Glial subtrees.
而 The Immune subtree was partitioned into myeloid and lymphoid lineages.
肿瘤内的基质细胞有多重层级结构
前面说到了很多文章是抓住stromal 里面的 fibro 和endo进行细分,并且编造生物学故事的。这个是肿瘤微环境(Tumor Microenvironment, TME)的重要组成部分,它们与肿瘤细胞相互作用,影响肿瘤的生长、侵袭和转移。基质细胞主要包括以下几种类型:
- 成纤维细胞(Fibroblasts):在肿瘤微环境中,成纤维细胞可以分化为癌症相关成纤维细胞(Cancer-Associated Fibroblasts, CAFs),它们通过分泌多种细胞因子和细胞外基质(ECM)成分,促进肿瘤的发展和转移。
- 内皮细胞(Endothelial Cells):内皮细胞在肿瘤血管生成中起到关键作用,它们可以形成新的血管,为肿瘤提供氧气和营养。
- 平滑肌细胞(Smooth Muscle Cells, SMCs):在某些情况下,平滑肌细胞也存在于肿瘤微环境中,尤其是在血管结构中。
- 周细胞(Pericytes):周细胞是血管壁的一部分,它们与内皮细胞相互作用,参与血管的稳定和成熟。
这些基质细胞通过分泌生长因子、细胞因子、趋化因子和代谢产物,以及通过细胞外基质的重塑,共同构建了一个复杂的肿瘤微环境,对肿瘤的生物学行为产生重要影响。了解这些细胞类型及其功能对于癌症的治疗策略开发至关重要。除去成纤维细胞之后的 Endothelial cells (ECs) and mural cells (MCs) are primary vasculature components that are directly involved in tumour angio-genesis 。其中 mural cells (MCs) 壁细胞包括周细胞(Pericytes)和平滑肌细胞(Smooth Muscle Cells, SMCs),它们位于内皮细胞层的外部,嵌入在血管的基底膜中。
它们这4大细胞亚群有时候界限不清楚,会互相掺杂,而且好难说清楚到底是该细分到什么程度算是一个头。
指数
2024新鲜出炉的论文(单细胞分辨率的肿瘤脉管图谱),英文题目为《Tumor vasculature at single-cell resolution》,论文通讯作者为重庆大学附属三峡医院副院长印明柱教授。重庆大学附属三峡医院作为第一完成单位和第一通讯作者单位联合清华大学、北京协和医院以及中南大学湘雅医院。可以看到里面的有单细胞的10x和bd这两个国外公司,还以寻因和新格元这两个国产单细胞公司的产品,来自于 31 种癌症类型的 372 个供体的大约 200,000 个细胞,主要是内皮细胞。其中 mural cells (MCs) 壁细胞包括周细胞(Pericytes)和平滑肌细胞(Smooth Muscle Cells, SMCs),如下所示就没有清晰的界限:

那么研究者是如何确定这两个单细胞亚群是否应该是细分呢?文章写到:
We also noted lower cell purity in PCs compared with SMCs (Extended Data Fig. 7b), prompting the heterogeneous phenotypes of PCs.
也就是说,作者把周细胞细分成为了4个亚群,如下所示:
- (1) vasodilation-associated PCs (vdPCs);
- (2) adipocyte-like PCs;
- (3) myoid-like PCs;
- (4) matrix-producing PCs (matPCs)
但是没有对平滑肌细胞继续细分,因为cell purity问题,让我们看看(Extended Data Fig. 7b), 是一个简单的箱线图:
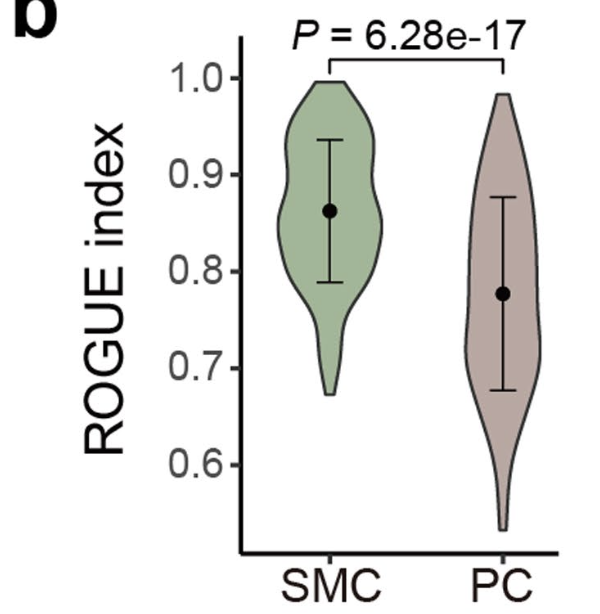
b. Violin plot showing cell purity for SMC (n = 10,819) and PC (n = 29,572) by ROGUE.
如何计算ROGUE index呢
有一个r包可以完成,大家可以安装它并且读一下它的官网说明书 :
# devtools::install_github("PaulingLiu/ROGUE")
# https://github.com/PaulingLiu/ROGUE/blob/master/vignettes/ROGUE_Tutorials.html
仍然是使用大家熟悉的数据集作为案例:
library(SeuratData) #加载seurat数据集
library(Seurat)
getOption('timeout')
options(timeout=10000)
# InstallData("pbmc3k")
data("pbmc3k")
sce <- UpdateSeuratObject(pbmc3k)
table(sce$seurat_annotations)
library(tidyverse)
suppressMessages(library(ROGUE))
expr <- GetAssayData(sce , slot="counts") %>% as.matrix()
expr <- matr.filter(expr, min.cells = 10 )
ent.res <- SE_fun(expr)
SEplot(ent.res)
rogue.value <- CalculateRogue(ent.res, platform = "UMI")
可以看到,这个表达量矩阵是来源于pbmc里面是各种各样的免疫细胞,所以可以继续细分,所以它的ROGUE index是很小的:
> rogue.value
[1] 0.7476979
# ROGUE范围为0~1,ROGUE = 1表面该亚群纯度很高;
# 相反ROGUE ~0,表面该亚群纯度较低可以被继续细分
# ROGUE评分默认>0.9时是一致性比较高的细胞亚类
我们有两个方法可以看看具体的每个单细胞亚群的ROGUE index
rogue.res <- rogue(expr, labels = sce$seurat_annotations ,
samples = sce$orig.ident,
platform = "UMI", span = 0.6)
rogue.res
rogue.boxplot(rogue.res )
> as.data.frame(t(rogue.res))
pbmc3k
Memory.CD4.T 0.9727606
B 0.9766577
CD14..Mono 0.9717287
NK 0.9666515
CD8.T 0.9707980
Naive.CD4.T 0.9819104
FCGR3A..Mono 0.9656822
NA. NA
DC 0.9424454
Platelet 0.9762869
可以看到的如果是前面的2700个pbmc单细胞分群后在每个单细胞亚群内部其实ROGUE index都提升了,之前的0.74,现在普遍是0.95以上,因为本来pbmc就是混合体嘛,这样的话细分亚群是合理的 ,但是我如果是自己把这个 2700个pbmc单细胞 表达量矩阵拆分后独立走 CalculateRogue 函数计算 ,发现其实是每个亚群仍然是有提升的空间 :
sort(table(sce$seurat_annotations))
y=unique(sce$seurat_annotations ) ;y
z=lapply(y[1:7], function(x){
#x=y[1];x
CalculateRogue(SE_fun(matr.filter(expr[,sce$seurat_annotations == x], min.cells = 10 )),
platform = "UMI")
})
> data.frame(cell=y[1:7],index=unlist(z))
cell index
1 Memory CD4 T 0.8563656
2 B 0.8480198
3 CD14+ Mono 0.8512200
4 NK 0.7976365
5 CD8 T 0.8116135
6 Naive CD4 T 0.8945871
7 FCGR3A+ Mono 0.7220867
略微有点尴尬啊, 我没有看这个ROGUE index算法,但是感觉不稳定?